
- ÍNDICE -
1. Definición y funciones principales
2. Composición
3. Propiedades físicas
4. Glucocálix
5. Transporte de moléculas pequeñas
6. Transporte de moléculas grandes
7. Superficie celular
4. Glucocálix
5. Transporte de moléculas pequeñas
6. Transporte de moléculas grandes
7. Superficie celular
1. DEFINICIÓN Y FUNCIONES
La membrana plasmática es un
componente celular con diversas funciones, como la de aislar y proteger el
interior celular, permitir intercambiar sustancias de una forma regulada e
interaccionar con otras células.
 |
| Las membranas de las células están compuestas, en orden de abundancia, de lípidos, proteínas y glúcidos |
2. COMPOSICIÓN DE LA MEMBRANA
PLASMÁTICA
Está compuesta por una capa
doble (bicapa) de lípidos (componente más abundante) , proteínas y algunos
glúcidos.
A) LÍPIDOS
En cuanto a los lípidos que la
componen, podríamos decir que son, principalmente:

1. Fosfolípidos: Son el ácido
graso mayoritario. Tienen una parte hidrófila y otra hidrófoba. Esto condiciona
la morfología de los agrupaciones de fosfolípidos, que tienden a disminuir las
partes hidrófobas en contacto con el agua. Debido a la morfología cilíndrica de
un fosfolípido, esta agrupación es la bicapa. Algunos tipos de fosfolípidos son
las cefalinas, lecitinasd o la Fosfatidilserina, la única con una cabeza polar
negativa, responsable de que la carga en el interior de la célula sea negativa
con respecto al exterior. Los fosfolípidos
no pueden pasar de una bicapa a otra de la membrana salvo circunstancias
especiales (por ejemplo, unidos a una traslocasa de lípidos), pero sí pueden
hacer desplazamientos laterales – que son muy rápidos: pude llegar a haber
hasta $10^7$ difusiones laterales en un solo segundo -.
2. Esfingolípidos. Un ejemplo es
la esfingomielina, presente en las neuronas e implicada en la velocidad de la
transmisión del impulso nervioso.
3. Glucolípidos. Poco abundantes
pero esenciales ya que están implicados en el reconocimiento celular.
4. Colesterol. Aproximadamente
el 35% de todos los lípidos que componen la membrana y, en relación a los
cuales, es de un tamaño pequeño.
B) PROTEÍNAS
Las proteínas también desempeñan
funciones muy variadas, como la enzimática, la estructural, la de transporte o
la de recepción de información (se verá más adelante). Podemos encontrarnos
proteínas integrales (completamente insertas en la membrana, de forma que
necesitamos detergentea para separarlas) o proteínas periféricas (con una unión
mucho más débil a la membrana).
 |
| Izquierda: comparación entre una proteína integral de membrana en alfa hélice (como en los canales o las bombas celulares) y en beta-lámina (permite la entrada considerable de agua en mitocondrias, células vegetales...) |
1. Proteínas integrales. Pueden tener tanto estructura alfa hélice
como lámina beta. Las proteínas de alfa hélice tienen la parte hidrófoba hacia
el interior de la membrana. Los canales de las células son de este tipo. Sin
embargo, una proteína en beta-lámita forma unas estructuras de barril (aunque las
partes hidrófilas siguen quedando hacia el exterior), que permiten la entrada
masiva de agua y las sustancias que ésta contenga. Están presentes en
mitocondrias, células vegetales, cloroplastos y bacterias.
2. Proteína periféricas. Pueden
anclarse a la membrana de muchas formas: covalentemente, iónicamente, asociándose
a proteínas integrales de membrana o insertándose parcialmente en la membrana.
Hay un ejemplo especial, las proteínas unidas de forma covalente con cola GPI,
que conforman los antígenos de superficie.
Al igual que ocurría en los fosfolípidos, las proteínas pueden
trasladarse e incluso rotar, pero no pueden pasarse de una bicapa a otra.
Aunque presentan cierto dinamismo, cuando las proteínas se encuentran asociadas
a las proteínas del citoesqueleto o formando desmosomas (uniones de células muy
fuertes, que impiden el paso de sustancias), permanecen quietas.
3. PROPIEDADES FÍSICAS
Las
propiedades físicas dependen de los lípidos.
Las
propiedades de la membrana son:

a) Fluidez. Es de suma importancia,
pues algunos procesos de transporte y actividades enzimáticas pueden detenerse
cuando la viscosidad de la bicapa se incrementa más allá de un nivel umbral. La
fluidez depende de tres factores: saturación, temperatura y del colesterol. En
cuanto a la saturación: Si hay alguna instauración en el ácido graso, el
empaquetamiento entre fosfolípidos es menor y, por lo tanto, hay mayor fluidez.
Además, una menor longitud en la cadena reduce la tendencia de las colas
hidrocarbonadas a interaccionar entre sí. En cuanto a la temperatura: A mayor
temperatura, mayor fluidez. Y, por último, en cuanto al colesterol: El
colesterol se inserta en la bicapa de forma que su región más rígida y plana
interactúa con – y en parte inmoviliza –parte de los fosfolípidos. Por lo
tanto, disminuye su movilidad, incrementando su rigidez y reduciendo su permeabilidad
a pequeñas moléculas solubles en agua. Además, a bajas temperaturas, el
colesterol impide que se solidifique la membrana.
b) Asimetría. Los
fosfolípidos no se distribuyen igual en ambas caras, como también ocurre con
las proteínas; es decir, los fosfolípidos presentes en la cara interna de la
membrana son diferentes a los que aparecen en la cara externa. Un ejemplo de
esto es la Fosfatidilserina, que sólo se muestra en la membrana interna (sólo
se muestra en la externa si la célula va a sufrir apoptosis). Por otra parte,
la esfingomielina suele aparecer en la cara externa y, además, forma las balsas
lipídicas. El colesterol puede estar en ambas regiones, y es la única molécula
lipídica que puede hacer un movimiento de flip-flop (pasarse de una bicapa a
otra). Los lípidos que tienen restos glucídicos (glucolípidos), se muestran en
la cara exterior.
La
asimetría permite diferencias de carga eléctrica entre el medio intracelular y
el extracelular (potencial de membrana. En el interior el potencial es
negativo, puesto que está la F. Serina), permite el reconocimiento celular y la
señalización de células muertas (pues, como dijimos, la Fosfatidilserina sólo
aparece en la cara externa cuando la célula está sufriendo apopotosis), etc.
c) Heterogeneidad. La distribución de los componentes dentro de la célula es distinta; están localizados en zonas concretas, como los lipid rafts, en los cuales se acumula un fosfolípido concreto, e incluso colesterol. Son rígidas, tienen mayor grosor (debido a que las cadenas de los esfingolípidos de las mismas son más largas y rectas que las del resto de lípidos de membrana, lo que permite acomodar mejor ciertas proteínas) y se mantienen unidos por fuerzas de Van der Waals. En ellas se encuentran determinadas proteínas que viajan juntas (se agrupan porque deben estar próximas entre sí para su correcto funcionamiento).


 |
| Las proteínas - y fosfolípidos - presentes en las diferentes bicapas de la membrana no son iguales. SM=Esfingomielina. PS=Fosfatidilserina. PC= Fosfatidilcolina |
c) Heterogeneidad. La distribución de los componentes dentro de la célula es distinta; están localizados en zonas concretas, como los lipid rafts, en los cuales se acumula un fosfolípido concreto, e incluso colesterol. Son rígidas, tienen mayor grosor (debido a que las cadenas de los esfingolípidos de las mismas son más largas y rectas que las del resto de lípidos de membrana, lo que permite acomodar mejor ciertas proteínas) y se mantienen unidos por fuerzas de Van der Waals. En ellas se encuentran determinadas proteínas que viajan juntas (se agrupan porque deben estar próximas entre sí para su correcto funcionamiento).
4. GLUCOCÁLIX
El glucocálix es una región de
glúcidos que recubre las células, y sus funciones son la protección o la de
facilitar la adhesión.
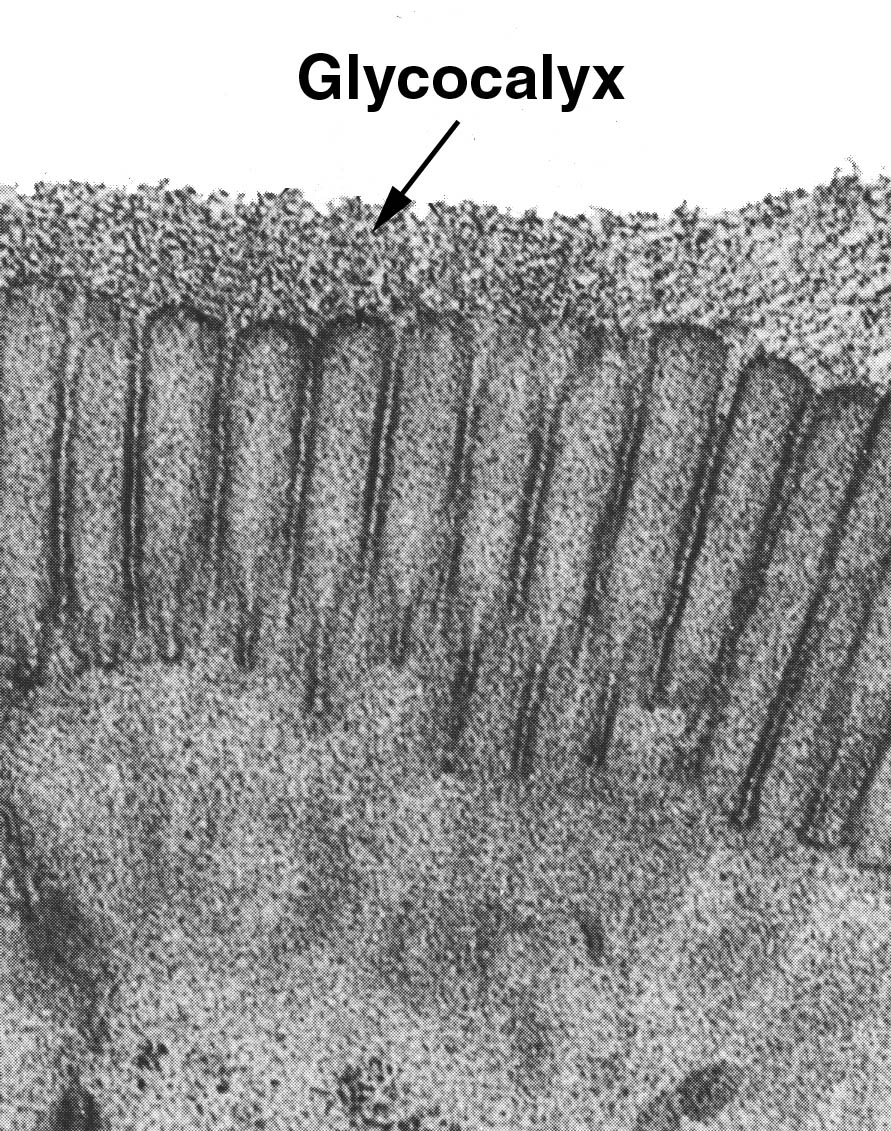 |
| El glucocálix es una región de la membrana plasmática externa recubierta de glúcidos con diversas funciones [Fuente] |
5. TRANSPORTE A TRAVÉS DE MEMBRANA (MOLÉCULAS PEQUEÑAS)
Las sustancias pueden atravesar
la membrana por sí mismos o con ayuda. En realidad, todas las sustancias,
pasado un determinado tiempo que puede variar mucho dependiendo de las
características de las mismas (hidrofobia, tamaño, carga…), terminarían
atravesando la membrana. Las sustancias que están libres en el citoplasma (moléculas
pequeñas y apolares, como el oxígeno, el nitrógeno o el dióxido de carbono u
moléculas apolares como las hormonas esteroideas) a veces consiguen
atravesarlas por sí mismas (difusión simple). El agua, al igual que la urea,
también es capaz de atravesar la membrana, aunque este proceso es más lento.
Por otra parte, las moléculas más grandes, necesitan facilidades para el
transporte, puesto que son incapaces de atravesar por sí mismas la membrana.
 |
| Muchas moléculas, debido a su pequeño tamaño, pueden atravesar la membrana plasmática por sí solas |
Es recomendable aclarar lo que
es el gradiente. Éste implica la diferencia de una magnitud en diferentes
regiones del espacio. Por ejemplo, que haya una temperatura diferente en el aire cercano a un
radiador encendido respecto a un lugar alejado a él, supone una diferencia de temperatura,
un gradiente de temperatura, que lleva asociado el movimiento de las partículas
de aire con el fin de homogeneizarlo todo. También pueden ser provocado por el desequilibrio de otras dos magnitudes:
a) Gradiente de concentración.
Que en una zona haya mayor concentración de una determinada sustancia que en la
otra. Ósmosis.
b) Gradiente eléctrico, debido a
la carga; es decir, a los iones que se encuentran en ambos medios celulares.
Cuando se tienen en cuenta tanto
el de concentración como el eléctrico (en moléculas con carga), el gradiente
resultante es el electroquímico.
 |
| Ejemplo del gradiente de concentración |
Por lo tanto, podemos
diferenciar tres formas de atravesar la membrana:
1. Difusión Simple. Ésta no sucede para igualar gradientes entre el
medio extracelular y el intracelular, sino que se debe a la probabilidad (si
hay más concentración de cierta molécula en el medio extracelular, por ejemplo,
es más probable que estas moléculas entren a que salgan). Por esto se dice que
siempre la difusión simple va a ser a favor de gradiente (por lo que no va a
requerir gasto de energía). La difusión simple, en conclusión, no requiere
transportadores ni es selectiva.
2. Transporte Pasivo. Puede deberse a canales o a proteasas. Sin gasto de energía y a favor de gradiente.
a) Proteínas canal. Estas
proteínas tienen tres características: son muy específicas (sólo ión cloruro, o
ión sodio, etc.), dan lugar a un transporte muy rápido (siendo esto esencial
para ciertos procesos) y no están siempre abiertos. Algunos ejemplos de canales
son:
- Canales de Agua. Se les
llama acuaporinas, y sólo permiten el paso de agua debido a que poseen ciertos
restos internos. Sin embargo, estos canales están siempre abiertos. Su
estructura es hélice-alfa y son muy importantes en el riñón (nefronas) y en las
plantas
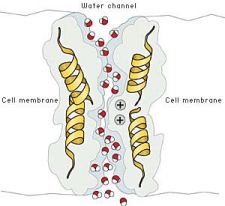
- Canales de Sodio. Los
iones en disolución están asociados a las moléculas de agua. Una molécula Na+
- H2O puede pasar perfectamente por este canal, pero la
asociación K+-H2O es demasiado grande; no
cabe.
- Canales TRP. Se abren
por la diferencia de temperatura. Actúan como receptores de temperatura (por
ejemplo, el canal CMR1 se abre con el metanol, y l VRL-1, con la capsaicina del
chile picante).
Como se dijo antes, los canales
no están siempre abiertos, sino que pueden estar cerrados e, incluso,
inhibidos. Los que se abren y cierras pueden depender, o bien del ligando (ej.: sinapsis -> Cuando el
neurotransmisor se liga a un lugar específico, el canal se abre), o bien de la temperatura (como los recién
descubiertos TRP), o bien del voltaje
(ej.: canal de K+ -> Cuando se invierte el potencial de membrana
[región intracelular positiva; extracelular, negativa], se abre este canal, de
forma que los iones K+ salen). Por otra parte, los canales pueden
ser inhibidos por toxinas, como ocurre con medicamentos contra la acidez
estomacal (omeprazol, almax…), pues bloquean los canales de Cl-.
b) Permeasas (Transportadores de
membrana). Transportan moléculas grandes sin carga, como la glucosa (azúcares,
aminoácidos y nucleótidos). El soluto se une a un lugar específico de la
permeasa. Ésta cambia su conformación, de forma que disminuye su afinidad con
el sustrato y la suelta (ya en el medio intracelular).
 |
3. Transporte Activo. Sucede en contra de gradiente de
concentración y con gasto de energía (ATP, luz …). Se gasta mucha energía para
mantener el potencial de membrana, para lo que ha de controlarse la
concentración absoluta de las diferentes sustancia
En un medio hipotónico, para
evitar la plasmólisis, las células vegetales han desarrollado la pared vegetal
[rígida, difícilmente deformable]; los protozoos acumulan el agua en vacuolas
que luego expulsan y las células animales sueltan hacia el exterior iones (lo
cual se realza en contra de concentración y, por lo tanto, requiere energía).

Al mismo tiempo, hay varios
tipos de transporte activo:
a) Bombas. Utilizan principalmente
ATP y luz. Podemos encontrarnos diferentes tipos:
a.1) Bombas F0/F1: La F0 es un
complejo proteico integral localizado en la membrana interna mitocondrial o del
cloroplasto. Cuando recibe H+, los desvía (la dirección a la que los
envía no es la misma de la que proceden), de forma que una proteína rota en uno o en otro sentido, permitiendo sintetizar
o hidrolizar al ATP.
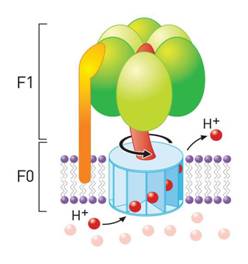 |
| Bomba del tipo F0-F1, capaz de sintetizar o hidrolizar ATP, dependiendo del sentido en el que rote la proteína [Fuente] |
a.2) Bombas ATPasas: El ejemplo por excelencia de esta familia es
la bomba de Na+/K+, que introduce dos iones potasio en la célula y saca al
exterior 3 de sodio (dicho grosso modo: hace el exterior más positivo y el
interior más negativo). En el medio extracelular hay una concentración de iones
de sodio más elevada que en el medio intracelular, por lo que va en contra del
gradiente de concentración (lo cual implica consumo de ATP).
 |
| Bomba de sodio-potasio [Fuente] |
a.3) Transportadores ABC: Estas bombas transportan moléculas
grandes, iones… Ejemplo: transportador CFTR: Proteína implicada en la fibrosis
quística. Si hay una mutación en esta
proteína, apenas sale ión cloruro al exterior y, por lo tanto, tampoco sale agua,
por lo que el moco es muy poco fluido, lo cual da lugar a la fibrosis quística.
b) Portadores. También llamados carriers. Puede ser:
- Uniporter (sólo se transporta una molécula y su velocidad viene
dada por la concentración de dicha molécula en el medio)
 |
| El transporte activo se hace desde donde hay menor concentración hacia donde hay mayor concentración (en contra de gradiente) y con gasto de energía (ATP, como se ve en la imagen) [Fuente] |
- Simporter (dos moléculas en el mismo sentido -> Ejemplo: Na+/Glucosa.
Introduce la glucosa en la célula transportando simultáneamente Na+).
Por otra parte, la localización
de las bombas y los portadores es de suma importancia. El complejo de unión
entre células impide que un compuesto pueda trasladarse de la zona apical hacia
la basal.
- Antiporter (dos moléculas en sentido contrario -> Na+/Ca2+)
En resumen, podemos concluir que las formas de transportar moléculas pequeñas son muy variadas, pudiendo clasificarse como:
6. TRANSPORTE A TRAVÉS DE MEMBRANA (MOLÉCULAS GRANDES)
6.1. ENDOCITOSIS
Podemos distinguir dos formas principales de transporte de moléculas de tamaño grande a través de la membrana plasmática (endocitosis): pinocitosis (moléculas relativamente pequeñas) y fagocitosis (Estructuras muy grandes, como bacterias, virus o cuerpos extraños). La pinocitosis, a su vez, puede ser de varios tipos: macropinocitosis (introducción de gran cantidad de agua), por vesículas recubiertas de clatrina, vesículas desnudas o por vesículas recubiertas de caveolina. Todas las vesículas se forman por la invaginación de la membrana.
Podemos distinguir dos formas principales de transporte de moléculas de tamaño grande a través de la membrana plasmática (endocitosis): pinocitosis (moléculas relativamente pequeñas) y fagocitosis (Estructuras muy grandes, como bacterias, virus o cuerpos extraños). La pinocitosis, a su vez, puede ser de varios tipos: macropinocitosis (introducción de gran cantidad de agua), por vesículas recubiertas de clatrina, vesículas desnudas o por vesículas recubiertas de caveolina. Todas las vesículas se forman por la invaginación de la membrana.

1. Pinocitosis.
1.1.
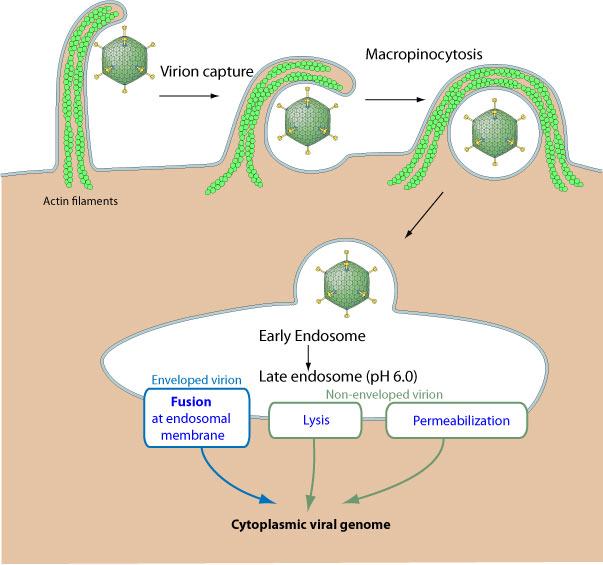
Macropinocitosis. La célula proyecta parte de su membrana hacia el exterior
formando un brazo que se va a fusionar con otra zona cercana de la membrana
plasmática. La formación de este brazo se debe a la polimerización de actina
 |
| Proyección de un brazo de actina para incluir endocitar un virus. Macropinocitosis. [Fuente] |
1.2.
Vesículas recubiertas por clatrina. La introducción de una sustancia en la
vesícula es muy específica, pues está mediada por receptores (que son proteínas
integrales de membrana, de las que hablamos previamente). Los receptores aparecen
en la membrana plasmática. La clatrina es una proteína que tiene una
conformación peculiar, pues está formada por unidades llamadas triskelion que, unidas forman una estructura de
red o canasta.
 |
| Molécula de clatrina, formado por trsikelion (azul clarito) [Fuente] |
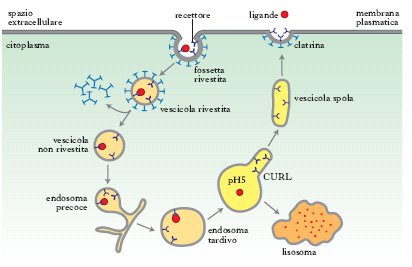
En el endosoma, por otra parte –que es un compartimento
membranoso (no es un orgánulo), intermediario entre el aparato de Golgi y el
lisosoma – se unen todas las vesículas de la endocitosis
mediada por clatrina. Como en su interior el pH es ácido, se produce la
liberación del receptor y del ligando provenientes de la vesícula. Este
endosoma temprano, con los contenidos de la vesícula en su interior, se va
acidificando por la acción de bombas de protones para finalmente llegar, mediante la fusión de
varios endosomas, a un lisosoma, donde el pH es muy bajo (aprox. 4.5), lo que
permite la degradación de los compuestos que habían llegado por la vesícula (así,
por ejemplo, queda el colesterol libre para la célula).
 |
| Producción de una vesícula recubierta por clatrina [Fuente] |
Los receptores se
reciclan en la membrana plasmática soltándose como vesículas, las cuales viajan
gracias a proteínas motoras que van sobre carriles normalmente de microtúbulos.
 |
| Molécula kinesina "caminando" sobre un microtúbulo mientras transporta una vesícula |
1.3. Vesículas recubiertas por caveolina (Caveolas). Éstas siempre
están formadas y aparecen especialmente en el endotelio. Se encargan, por
ejemplo, del transporte de HDL. En ellas, al igual que en las lipid rafts (balsas
lipídicas), los esfingolípidos y el colesterol son muy numerosos, pero se
diferencian de éstas en que poseen dos subunidades de caveolina. Estas
vesículas, al contrario de las recubiertas por clatrina, no van al endosoma,
sino al caveosoma (en su interior, el pH no es ácido y, además, es de menor
tamaño que en endosoma).
 |
| Tipos de endocitosis [Fuente] |
2. Fagocitosis. Introducción de grandes moléculas y cuerpos.
En este proceso, la actina comienza a polimerizar, engloba al cuerpo hasta que
forma una vesícula llamada fagosoma. A éste se le unen lisosomas y vesículas
del endosoma, recibiendo entonces el nombre de fagolisosoma, donde se degrada
el cuerpo que se halla en su interior.
 |
6.2. TRANSCITOSIS. No se trata de un proceso de introducción
o extracción de sustancias a través de la membrana plasmática, sino del envío
de sustancias de una parte de la célula hacia otras. Los marcadores y las
proteínas de una región de la membrana se encuentran inmovilizados, de forma
que para enviarlos de un extremo a otro de la célula, cruzando la misma, se
utilizan vesículas de transporte. Es decir, se produce una invaginación en la
región de la membrana plasmática donde se encuentra el receptor para englobarlo
en una vesícula que ya puede enviarse al destino requerido.
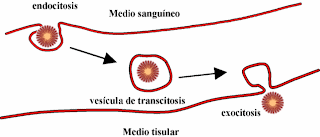 |
| La transcitosis es el transporte de una sustancia desde una zona de la célula a otra mediante vesículas [Fuente] |
6.3. EXOCITOSIS.
Expulsión de compuestos intracelulares. Las
vesículas proceden del aparato de Golgi y. Las proteínas encargadas de la
fusión entre las vesículas y la membrana plasmática son las SNARE. Cuando se aproximan entre sí, comienzan a
enrollarse de forma que acercan la vesícula hacia la membrana hasta que se
produce la fusión de las bicapas lipídicas. Algunas proteínas SNARE se
recuperan y reciclan por endocitosis.
Hay dos tipos:
4.1. Exocitosis Constitutiva. Sucede continuamente, como por
ejemplo la síntesis de lípidos y proteínas de membrana
4.2. Exocitosis Regulada. Productos de secreción específicos, como
las sustancias que componen la mucosa del tracto respiratorio.
7. SUPERFICIE CELULAR
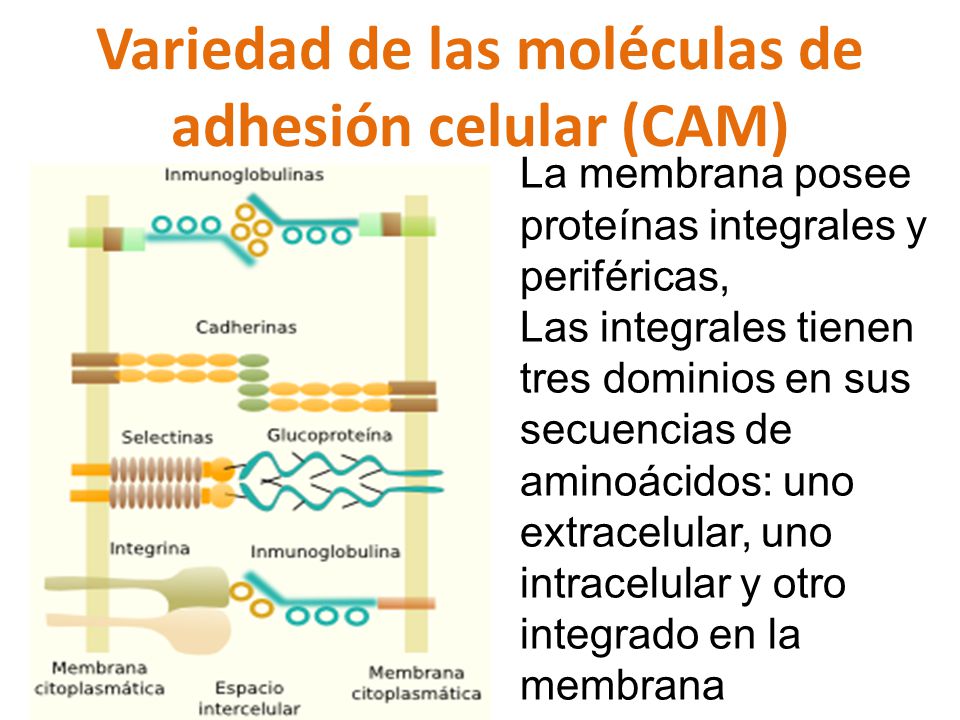
1. Composición. En la membrana plasmática se encuentran moléculas
de adhesión celular que permiten la unión de una célula con otra célula o con
la matriz extracelular. Algunas son la cadherina, selectina, integrina.
a) Cadherinas. Aparece por ejemplo en las sinapsis y, especialmente, en embriones. De hecho, a
medida que éste se desarrolla, este tipo de uniones, inicialmente tan
numerosas, van siendo reemplazadas por otro tipo.
b) Selectinas. Proteínas
con afinidad hacia restos azucarados. Aparece en el endotelio, en células
sanguíneas (permite la diapédesis) y en las plaquetas (favorece la coagulación
sanguínea).
c) Integrinas.
2. Uniones intercelulares. En el tejido epitelial es en el que más
uniones intercelulares hay. Esto se debe a que una de sus funciones principales
es la de revestir, impermeabilizar y proteger.
1.1. Uniones Ocluyentes. Las
células están unidas fuertemente entre sí, formando redes e impidiendo el paso
de sustancias, incluso de agua, entre las propias células. Cuando hay varias
así unidas, forman una zona ocludens.
1.2. Uniones Adherentes.
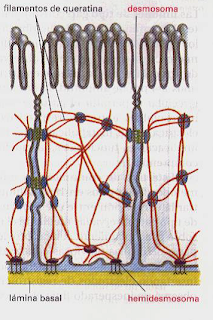
a) Desmosomas en banda.
b) Desmosomas puntuales. Tienen una distribución más dispersa que
los desmosomas en banda.
c) Hemidesmosomas. Son desmosomas que unen las células epiteliales
con la matriz extracelular. Están unidos en la parte interior a filamentos de
queratina y, en la parte exterior, con la lámina basal.
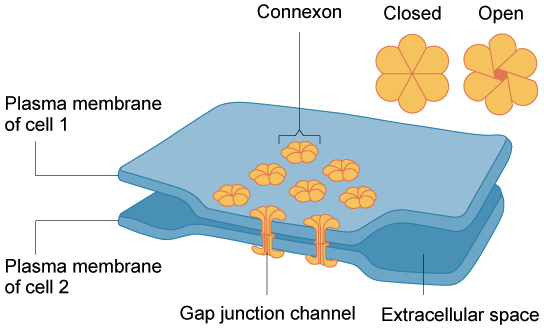
1.3. Uniones Comunicantes.
Permiten el paso de diferentes sustancias. Además, estas uniones están formadas
por pequeñas moléculas (conexinas) que se unen formando un poro (conexón) de tamaño relativamente
grande (1.5 nm)
3. Especialización (Diferenciación) de la membrana
Podemos encontrar, en tejidos
como el epitelial, donde hay polaridad (hay una diferencia morfolófica o de
composición entre la zona superior y la inferior), diferentes
especializaciones:

3.1. Apicales
a) Microvellosidades. Son
proyecciones de la membrana que aparecen en la región apical de la membrana y
que aumentan la superficie expuesta de la misma. Su conformación se mantiene
por filamentos de actina (la red de filamentos de actina se une con los
filamentos de los desmosomas en banda).
b) Estereocilios. Son
estructuras similares a las microvellosidades (es decir, son proyecciones de la
membrana) pero de mayor tamaño. Al ser tan grandes, se anastomosan (en algunas
regiones se fusionan). Su función es incrementar la superficie de absorción (en
el epidídimo –> testículos) e intervienen en el sentido del equilibrio (en
el oído).
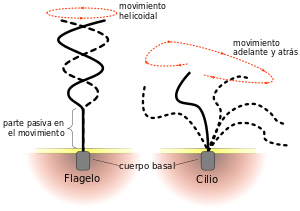
c) Cilios. Están
presentes en numerosos organismos unicelulares. Tienen una función motora. En
el aparato respiratorio se encargan del movimiento del moco, y en las trompas
de Falopio, del óvulo.
d) Flagelos. Son
similares a los cilios, pero tienen mayor tamaño y, además, su movimiento es de
rotación.
3.2. Laterales: Interdigitaciones
Aumentan la superficie de
contacto entre células (lo cual permite una mejor adhesión, puesto que hay más
espacio para desmosomas y uniones comunicantes).
3.3. Basales: Invaginaciones
Se incrementa la superficie de
contacto entre la célula epitelial y el tejido conjuntivo. Aparecen en tejidos
donde es muy necesario el intercambio de sustancias.
|







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
No hay comentarios:
Publicar un comentario